Week 3: Relabeling & Retraining
Welcome back to my blog! Last week, I illustrated the cerebellar basis of my project through outlining specifics on the cerebellar dentate nuclei, though this week, I’ll cover another aspect of this project: adolescence – a critical period in cognitive development.
While the cerebellum is associated with motor functions, recent studies have also demonstrated its role in cognitive functions, which is what I’ll be placing emphasis on in this project. Adolescence is considered a critical period in neurobiology as there are significant social developments and refinements in higher-order cognitive abilities, including working memory and cognitive control. Furthermore, critical periods are often triggered by both the reduction of the excitation/inhibition balance of neurotransmitters and experience, which encompasses biological, genetic, and experiential factors. As a result, critical periods shape the plasticity of cerebellar circuits, and it’s believed that this critical period opens at puberty as the mesocortical dopamine system begins developing throughout at this time (Larsen & Luna, 2018). Ultimately, cognitive abilities that are refined throughout adolescence establish their successive stability in adulthood, which is why certain perturbations throughout this critical period can actually lead to a lack of this stabilization later in life. Now onto analysis and my experience with SLEAPing this week!
As noted in my previous blogs, the key outcome measures that I’m using in this project include the time spent at the reward area, time spent at the digital touchscreen, and the time spent in differentiated behaviors. When I initially ran some of the Hierarchical Data Format 5 files (HDF5 files) on Google Colab, where we use Python for analysis, I noticed that the time spent at the digital touchscreen was fairly inaccurate as I was extracting points that averaged less than 5 seconds. Upon checking each video of the recorded mouse behavior, it was evident that I should have been generating data points that were far above that value as the mouse spent much more time interacting with the touchscreen. To rectify this issue, I began relabeling frames with the mouse at the digital touchscreen as the model likely wasn’t correctly identifying the nose1 node throughout those frames. I relabeled just under 300 new frames on our deep-learning software, SLEAP, for a total of 1,175 labeled frames. Once that was complete, I began retraining the model, which we’ll now call our n=1175 model. Below is the training graph for the n=1175 model.
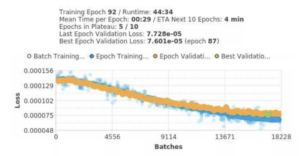
Specific metrics to note for the n=1175 model were 95.8% precision and 77.8% recall. Though the recall value is slightly under our target value of 80%, the model is predicting far better than the n=882 model, especially when the mouse is near the touchscreen. Unfortunately, I did have to scrap my older HDF5 files from the n=882 model, but luckily, it’s much easier to extract new HDF5 files with ASU’s supercomputer, Sol, rather than in my local environment.
Next week, I’ll continue with generating data from these newly-generated HDF5 files with Python. Other than my time spent SLEAPing this week, I was able to observe a brain tissue sectioning procedure with the cryostat. These will later be used for immunohistochemistry, which I’m hoping to start in the next couple of weeks to analyze c-Fos, an immediate early gene, in order to quantify neural activity. Stay tuned!
References:
Larsen B, Luna B. (2018) Adolescence as a neurobiological critical period for the development of higher-order cognition. Neuroscience & Biobehavioral Reviews. https://doi.org/10.1016/j.neubiorev.2018.09.005